Summary: Purkinje cells can modulate and filter incoming signals. The findings shed new light on learning mechanisms within the cerebellum.
Source: Kyoto University
In mere milliseconds trillions of chemical reactions ignite signals that travel across the billions of neurons in our brain. As we go through our daily lives and absorb new knowledge these neurons begin to modify themselves and change their signaling properties.
However, the mechanisms of how signals are integrated into the neurons to establish such flexibility, also known as plasticity, remains elusive.
Publishing in the Journal of Neuroscience, Gen Ohtsuki of Kyoto University’s Hakubi Center reports that Purkinje cells — the primary output neurons in the cerebellum — have the ability to modulate and filter incoming signals. The findings bring new insight into the learning mechanisms of the cerebellum and the brain.
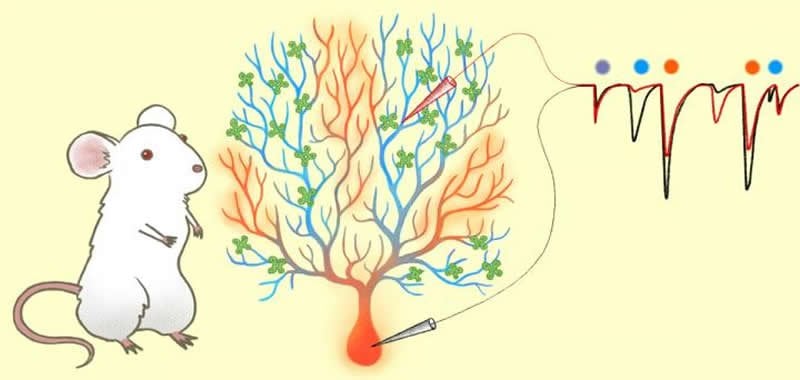
The cerebellum is a structure located at the base of the brain, and is known to play a vital role in motor control and cognitive function. Recent findings have even revealed its contributions in mental illnesses. One of the most vivid features of Purkinje cells are their long complex branches called dendrites.
It is thought that the plasticity of these Purkinje-cell dendrites is the basis for cerebellar learning. However, validation of this hypothesis was difficult due to the challenge of measuring signals within a single cell.
Thankfully in a prior study, Ohtsuki was successful in measuring the electrical activity on dendrites of a single Purkinje cell utilizing the patch-clamp method.
“To measure how electrical signals travel through the Purkinje cell membrane, I applied this method using rats and measured the spontaneous synaptic activity between the dendrite and the ‘soma’ or cell body,” explains Ohtsuki.
What he found was that signals coming from dendrites far away from the soma, known as distal dendrites, were not being registered. This suggests the dendrites have a mechanism that limits electroconduction, and that individual branches can choose whether an input passes through or not. In fact, the same signals were registered when they came from proximal dendrites — the ones closer to the soma.
After further analysis it was found that these distal dendrites modulated their incoming signals through intrinsic plasticity associated with the down-regulation of an ion channel called SK channels.
“One of the reasons for this new finding is because similar experiments used cesium ions in the intracellular fluid, so the phenomena itself could not be observed at all,” states Ohtsuki. “The results reveal a new learning mechanism at the dendritic level.”
He hopes to further verify these results and determine whether similar findings can be obtained with animals other than rodents, such as fishes and reptiles, or higher mammals.
Ohtsuki concludes, “Studying these fundamental processes should help us understand the reasons for the mechanism of intelligence.”
Source:
Kyoto University
Media Contacts:
Raymond Kunikane Terhune – Kyoto University
Image Source:
The image is credited to Kyoto University/Mindy Takamiya.
Original Research: Closed access
“Modification of synaptic-input clustering by intrinsic excitability plasticity on cerebellar Purkinje cell dendrites”. Gen Ohtsuki.
Journal of Neuroscience doi:10.1523/JNEUROSCI.3211-18.2019.
Abstract
Modification of synaptic-input clustering by intrinsic excitability plasticity on cerebellar Purkinje cell dendrites
The role of dendrites in the integration of widespread synaptic activity has been studied in experiments and theories (Johnston et al., 1996; Magee, 2007). However, whether the conduction of synaptic currents from dendrites to the soma depends on excitability of those dendritic branches is unclear. How modulation of the branch excitability affects the conduction of synaptic inputs and their selection on dendrites is also elusive. Here, I performed simultaneous voltage-clamp recordings from the soma and dendrites of single cerebellar Purkinje neurons in male Sprague-Dawley rats and analyzed the relationship between spontaneous excitatory postsynaptic currents (EPSCs) on both sides. I found that EPSCs on distal dendrites have a salient discordance in amplitude, compared to those on the soma. Furthermore, individual ratios of the EPSC concurrently recorded on the soma and dendrites were not unique, but discrete, suggesting the occurrence of various attenuations in different paths of dendritic branches to the soma. The obtained data and simulations indicate several distinct groups (4.5 ± 0.3, n = 22 somato-dendritic recordings) of co-occurred synaptic inputs in Purkinje cell dendrites. This clustering of synaptic currents was suggested to emerge at farther distances than the secondary bifurcations. Finally, ratios of the co-EPSCs were uniformly distributed after either intrinsic plasticity induction or SK-channel blockade. Overall, results suggest that in Purkinje cells the excitability along the dendrite processes modulates the conduction of EPSCs and makes active inputs heterogeneous through SK channel activity, intrinsic plasticity and dendritic branching. These properties of dendrites may confer branch-specific computational power to neurons.
SIGNIFICANCE STATEMENT
I have previously studied the “non-synaptic” plasticity of the intrinsic excitability in the cerebellar Purkinje cells (Belmeguenai et al., 2010), and branch-specific increase of intrinsic excitability of the dendrites (Ohtsuki et al., 2012; Ohtsuki and Hansel, 2018) through the downregulation of SK (small conductance Ca2+-activated K+) channels. In this study, I show that a dendritic filtering of synaptic electroconductivity is heterogeneous among the branches on distal dendrites and that the increase in the dendritic excitability accompanied with the intrinsic plasticity alters a state with the heterogeneity to a globally excitable state in Purkinje neurons. My findings propose a new learning model relying on the intrinsic excitability plasticity of the dendritic branch-fields.