

Summary: Researchers reveal how neurons set up and sustain the vital infrastructure that allows for seamless neurotransmission.
Source: Picower Institute for Learning and Memory
The nervous system works because neurons communicate across connections called synapses. They “talk” when calcium ions flow through channels into “active zones” that are loaded with vesicles carrying molecular messages.
The electrically charged calcium causes vesicles to “fuse” to the outer membrane of presynaptic neurons, releasing their communicative chemical cargo to the postsynaptic cell.
In a new study, scientists at The Picower Institute for Learning and Memory at MIT provide several revelations about how neurons set up and sustain this vital infrastructure.
“Calcium channels are the major determinant of calcium influx, which then triggers vesicle fusion, so it is a critical component of the engine on the presynaptic side that converts electrical signals to chemical synaptic transmission,” said Troy Littleton, senior author of the new study in eLife and Menicon Professor of Neuroscience in MIT’s Departments of Biology and Brain and Cognitive Sciences.
“How they accumulate at active zones was really unclear. Our study reveals clues into how active zones accumulate and regulate the abundance of calcium channels.”
Neuroscientists have wanted these clues. One reason is that understanding this process can help reveal how neurons change how they communicate, an ability called “plasticity” that underlies learning and memory and other important brain functions.
Another is that drugs such as gabapentin, which treats conditions as diverse as epilepsy, anxiety and nerve pain, binds a protein called alpha2delta that is closely associated with calcium channels. By revealing more about alpha2delta’s exact function, the study better explains what those treatments affect.
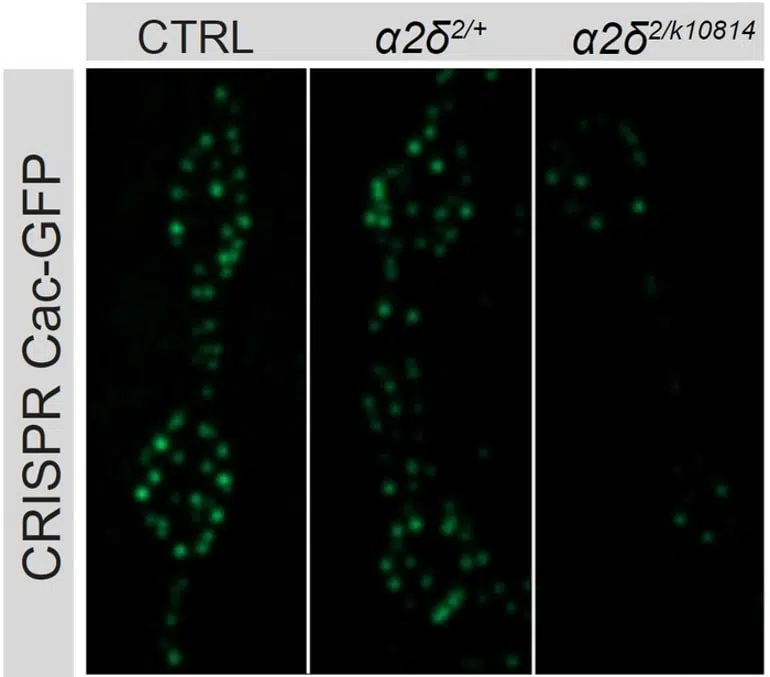
The more scientists knocked out a protein called alpha2delta with different manipulations (right two columns), the less Cac calcium channel accrued in synaptic active zones of a fly neuron (brightness and number of green dots) compared to unaltered controls (left column).
“Modulation of the function of presynaptic calcium channels is known to have very important clinical effects,” Littleton said. “Understanding the baseline of how these channels are regulated is really important.”
MIT postdoc Karen Cunningham led the study, which was her doctoral thesis work in Littleton’s lab. Using the model system of fruit fly motor neurons, she employed a wide variety of techniques and experiments to show for the first time the step by step process that accounts for the distribution and upkeep of calcium channels at active zones.
A cap on Cac
Cunningham’s first question was whether calcium channels are necessary for active zones to develop in larvae. The fly calcium channel gene (called “cacophony,” or Cac) is so important, flies literally can’t live without it. So rather than knocking out Cac across the fly, Cunningham used a technique to knock it out in just one population of neurons. By doing so, she was able to show that even without Cac, active zones grow and mature normally.
Using another technique that artificially prolongs the larval stage of the fly she was also able to see that given extra time the active zone will continue to build up its structure with a protein called BRP, but that Cac accumulation ceases after the normal six days.
Cunningham also found that moderate increases of decreases in the supply of available Cac in the neuron did not affect how much Cac ended up at each active zone. Even more curious, she found that while Cac amount did scale with each active zone’s size, it barely budged if she took away a lot of the BRP in the active zone. Indeed, for each active zone, the neuron seemed to enforce a consistent cap on the amount of Cac present.
“It was revealing that the neuron had very different rules for the structural proteins at the active zone like BRP that continued to accumulate over time, versus the calcium channel that was tightly regulated and had its abundance capped” Cunningham said.
Regular refresh
The team’s model shows factors that regulate Cac abundance at active zones. Active Zone scaffold development and Cac delivery via alpha2delta increases it while turnover keeps a lid on it. Cac biosynthesis barely increases aboundance.
The findings showed there must be factors other than Cac supply or changes in BRP that regulate Cac levels so tightly. Cunningham turned to alpha2delta. When she genetically manipulated how much of that was expressed, she found that alpha2delta levels directly determined how much Cac accumulated at active zones.
In further experiments, Cunningham was also able to show that alpha2delta’s ability to maintain Cac levels depended on the neuron’s overall Cac supply. That finding suggested that rather than controlling Cac amount at active zones by stabilizing it, alpha 2delta likely functioned upstream, during Cac trafficking, to supply and resupply Cac to active zones.
Cunningham used two different techniques to watch that resupply happen, producing measurements of its extent and its timing. She chose a moment after a few days of development to image active zones and measure Cac abundance to ascertain the landscape. The she bleached out that Cac fluorescence to erase it.
After 24 hours, she visualized Cac fluorescence anew to highlight only the new Cac that was delivered to active zones over that 24 hours. She saw that over that day there was Cac delivery across virtually all active zones, but that one day’s work was indeed only a fraction compared to what had built up over several days before.
Moreover, she could see that the larger active zones accrued more Cac than smaller ones. And in flies with mutated alpha2delta, there was very little new Cac delivery at all.
If Cac channels were indeed constantly being resupplied, then Cunningham wanted to know at what pace Cac channels are removed from active zones.
To determine that, she used a staining technology with a photoconvertible protein called Maple tagged to the Cac protein that allowed her to change the color with a flash of light at the time of her choosing. That way she could first see how much Cac accumulated by a certain time (shown in green) and then flash the light to turn that Cac red.
When she checked back five days later, about 30 percent of the red Cac had been replaced with new green Cac, suggesting 30 percent turnover. When she reduced Cac delivery levels by mutating alpha2 delta or reducing Cac biosynthesis, Cac turnover stopped. That means a significant amount of Cac is turned over each day at active zones and that the turnover is prompted by new Cac delivery.
Littleton said his lab is eager to build on these results. Now that the rules of calcium channel abundance and replenishment are clear, he wants to know how they differ when neurons undergo plasticity—for instance when new incoming information requires neurons to adjust their communication to scale up or down synaptic communication.
He said he is also eager to track individual calcium channels as they are made in the cell body and then move down the neural axon to the active zones, and he wants to determine what other genes may affect Cac abundance.
In addition to Cunningham and Littleton, the paper’s other authors are Chad Sauvola and Sara Tavana.
Funding: The National Institutes of Health and the JPB Foundation provided support for the research.
About this neuroscience research news
Author: David Orenstein
Source: Picower Institute for Learning and Memory
Contact: David Orenstein – Picower Institute for Learning and Memory
Image: The image is credited to Littleton Lab/MIT Picower Institute
Original Research: Open access.
“Regulation of presynaptic Ca2+ channel abundance at active zones through a balance of delivery and turnover” by Troy Littleton et al. eLife
Abstract
Regulation of presynaptic Ca2+ channel abundance at active zones through a balance of delivery and turnover
Voltage-gated Ca2+ channels (VGCCs) mediate Ca2+ influx to trigger neurotransmitter release at specialized presynaptic sites termed active zones (AZs). The abundance of VGCCs at AZs regulates neurotransmitter release probability (Pr), a key presynaptic determinant of synaptic strength. Although biosynthesis, delivery and recycling cooperate to establish AZ VGCC abundance, experimentally isolating these distinct regulatory processes has been difficult.
Here we describe how the AZ levels of Cacophony (Cac), the sole VGCC mediating synaptic transmission in Drosophila, are determined.
We also analyzed the relationship between Cac, the conserved VGCC regulatory subunit α2δ, and the core AZ scaffold protein Bruchpilot (BRP) in establishing a functional AZ. We find Cac and BRP are independently regulated at growing AZs, as Cac is dispensable for AZ formation and structural maturation, and BRP abundance is not limiting for Cac accumulation. Additionally, AZs stop accumulating Cac after an initial growth phase, whereas BRP levels continue to increase given extended developmental time. AZ Cac is also buffered against moderate increases or decreases in biosynthesis, whereas BRP lacks this buffering.
To probe mechanisms that determine AZ Cac abundance, intravital FRAP and Cac photoconversion were used to separately measure delivery and turnover at individual AZs over a multi-day period. Cac delivery occurs broadly across the AZ population, correlates with AZ size, and is rate-limited by α2δ.
Although Cac does not undergo significant lateral transfer between neighboring AZs over the course of development, Cac removal from AZs does occur and is promoted by new Cac delivery, generating a cap on Cac accumulation at mature AZs.
Together these findings reveal how Cac biosynthesis, synaptic delivery, and recycling set the abundance of VGCCs at individual AZs throughout synapse development and maintenance.




